ESTRUCTURA DEL COL�GENO
|
|
ESTRUCTURA DEL COL�GENO |
|
El col�geno es un material extracelular fabricado por los fibroblastos y es una prote�na fibrosa que resulta relativamente insoluble en agua, en contraposici�n a otras familias de llamadas globulares, que s� son solubles en agua. La base molecular del col�geno est� constituida por cadenas de polip�ptidos y cada uno de �stos es un pol�mero de amino�cidos. Es decir, son cadenas constituidas por amino�cidos, que son unidades moleculares peque�as. Cada uno de estos amino�cidos se caracterizan por tener por lo menos dos funciones distintas: una amino y una �cida en la misma unidad molecular. Los polip�ptidos no son m�s que cadenas de estos amino�cidos que se encuentran en los organismos biol�gicos en n�meros limitados. Existen unos 20 amino�cidos fundamentales. Su secuencia permite identificar las cadenas de polip�ptidos. Todas las prote�nas que existen en la biolog�a se identifican porque poseen una secuencia particular de amino�cidos que es �nica y propia de esa prote�na, que permite identificarla por m�todos bioqu�micos, inmunol�gicos, etc.
Estas macromol�culas de tropocol�geno son muy peque�as. S�lo se conocen por m�todos indirectos, son detectables bioqu�micamente . Las macromol�culas de tropocol�geno se agrupan entre s� constituyendo estructuras llamadas fibrillas de col�geno. Cada fibrilla de col�geno est� constituida por miles de mol�culas de tropocol�geno, que son visibles al microscopio electr�nico, se pueden detectar, medir, colorear, estudiar en forma relativamente c�moda. Si bien en algunas partes est�n aisladas, m�s o menos sueltas, en la mayor hurte del organismo, sobre todo en la dermis, centenares de estas fibrillas se unen lado a lado formando fibras col�genas mucho m�s voluminosas, visibles con microscopio �ptico. Las fibras col�genas tienden a agruparse en conjuntos m�s grandes llamados haces col�genos. Antes de entrar en m�s detalles de la constituci�n qu�mica de esta macromol�cula vamos a ver algunas propiedades f�sicas que son importantes. En primer lugar, el col�geno est� especialmente concentrado en aquellos tejidos que soportan peso (el peso del organismo), fundamentalmente los cart�lagos y los huesos. Tambi�n existe col�geno concentrado en altas proporciones en aquellas partes del organismo que transmiten fuerza, como los tendones (ligamentos que unen los m�sculos con las piezas esquel�ticas). En tercer lugar, el col�geno aparece en forma numerosa en aquellos lugares como la dermis o las fascias (l�minas que recubren los m�sculos) sirven pura proteger, o donde se necesita un material que resista la tracci�n o los cambios de volumen. Finalmente, el col�geno, en una de sus formas, constituye pr�cticamente una armaz�n de microfibrillas, que sostiene la estructura de todos los �rganos y v�sceras del organismo. 0 sea, que encontramos peque�as fibrillas de col�geno en el h�gado, en el bazo, en el pulm�n, en fin, no hay ninguna v�scera que no tenga esa armaz�n de col�geno. Por ejemplo los vasos sangu�neos que son tubos, mantienen su forma tubular en funci�n de que alrededor de la pared tienen una trama en espiral de fibrillas de col�geno, que constituyen el soporte de esa estructura tubular. En general, el col�geno aparece como un material altamente ordenado. En algunos lugares las fibras de col�geno se disponen en forma estrictamente paralela. El ejemplo m�s t�pico es el de los tendones. En otros lugares como la dermis, las fibras col�genas aparecen entrelazadas en todos los planos del espacio de un modo muy apretado. De modo que cuando nosotros observamos la dermis al microscopio �ptico, o sea con poca resoluci�n y sin ning�n artificio que nos permita separar las fibras de col�geno. pr�cticamente no podemos distinguir los l�mites entre una fibra y otra, porque est�n formando una malla demasiado apretada. Por eso la dermis vista al microscopio �ptico con coloraciones normales o de rutina aparece como un tejido conjuntivo casi homog�neo, donde de vez en cuando se ven c�lulas separadas. De ah� que cl�sicamente para estudiar el tejido conjuntivo de la dermis se hallan ideado una serle de m�todos para separar las fibras. Uno de los m�todos m�s cl�sicos es separar las fibras por m�todos mec�nicos y hacer una disociaci�n de las mismas con agujas, de modo de poder observarlas al microscopio con m�s comodidad. Pero el procedimiento tambi�n cl�sico pero m�s eficiente es producir un edema, es decir, inyectar la dermis con suero fisiol�gico de modo de aprovechar una de las propiedades de la sustancia fundamental amorfa que es una gran capacidad de hidratarse, de embeberse con agua y por lo tanto, aumentar el volumen y al hincharse la sustancia fundamental, las fibras col�genas aparecen m�s separadas En estas circunstancias, lo que vemos como material col�geno, son haces que tienden a veces a ramificarse, ondulados, que tienen dimensiones variables, poro que se miden en micras o en micr�metros, o sea en unidades que, est�n en el orden de 10 -6, o sea que cada micra es la mil�sima parte del mil�metro Los haces de fibras col�genas miden habitualmente entre 15 y 30 micras, aunque hay tambi�n haces m�s finos y m�s gruesos; en cada uno de estos haces, nosotros encontramos fibrillas, fibras de col�geno que se disponen paralelamente. Cuando el haz se ramifica, las fibrillas se distribuyen, pero las fibras en s� mismas no se ramifican. Es decir, lo que se puede ramificar es el haz, pero no las fibras de col�geno. Es frecuente encontrar en estos haces, en la visi�n del tejido conjuntivo, c�lulas, fibroblastos que est�n unidos, pegados a la superficie de los haces de col�geno. Esos fibroblastos o est�n sintetizando nuevo material col�geno. o est�n reponiendo material col�geno que se est� desgastando, porque una de las caracter�sticas de esta prote�na, igual que de muchas otras del organismo, es que una vez que se forman, despu�s que pasa el tiempo son sometidas a un constante recambio, o sea, hay partes que salen y partes que se renuevan. Otro concepto importante es el de que el col�geno forma parte de un complejo funcional que es el tejido conjuntivo.
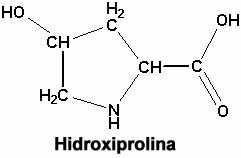
Cl�sicamente, para colorear los haces de fibras col�genas. con el microscopio �ptico, se han utilizado una variedad de colorantes que son �tiles para ponerlas en evidencia o para distinguirlas de otras fibras, como las musculares o mismo las el�sticas. As� por ejemplo existe un colorante llamado verde luz, que colorea selectivamente a las fibras col�genas. Del mismo modo, el azul de toluidina colorea m�s o menos selectivamente ir las fibras col�genas. Durante bastante tiempo se hablaba solamente de un tipo de fibras col�genas. Ya hace bastantes a�os, se comprob� que cuando se utilizaba un m�todo de coloraci�n basado en la impregnaci�n de tejidos con sales met�licas (en este caso la sal met�lica m�s utilizada ha sido el carbonato de plata) se pueden distinguir dos tipos de fibras col�genas. Un tipo de fibras que aparecen gruesas de un color rojo, y que abundan por ejemplo en la dermis, los tendones, en las c�psulas de los �rganos, etc. y fibrillas o fibras de col�geno m�s finas, que aparecen de color negro, que durante mucho tiempo se denominaron fibras de reticulina. 0 sea, que con m�todos de tinci�n al microscopio �ptico no podemos discriminar m�s que esos dos tipos de material col�geno. Cuando se utilizan m�todos bioqu�micos, inmunol�gicos y tambi�n la microscop�a electr�nica se ha llegado a discriminar en una primera etapa 5 tipos de fibras col�genas. y m�s modernamente, hasta 12. Vamos a referirnos en principio, a esos primeros 5 tipos, pero antes tenemos que hacer una breve incursi�n por la estructura ya m�s fina de estas fibras. Cuando estudiamos un peque�o fragmento del haz de fibras col�genas al microscopio �ptico, observamos un panorama bastante m�s complicado. Observamos fibrillas de un di�metro variable, pero que anda alrededor de las 100 milimicras, o sea, una unidad que a su vez es la mil�sima parte de la micra, por lo tanto es 10 -9 y esas fibras que tienen la longitud muy grande, a su vez aparecen subdivididas por una serie de bandas que en el microscopio electr�nico aparecen coloreadas de color oscuro o de color m�s claro y esas bandas se repiten a lo largo de la fibra constituyendo una. periodicidad (o sea, una estructura bandeada que se repite siempre igual a s� misma). La distancia entre las bandas que se repiten dentro de la fibrilla es siempre igual, entre 60 y 70 nan�metros. Habitualmente se habla de 64 nan�metros (esto es la periodicidad del col�geno). Y precisamente cuando se colocan estas fibras llamadas de col�geno nativo (o sea, col�geno que aparece naturalmente en los tejidos) en medio �cido, un medio con �cido ac�tico d�bil, se disocian, pierden la periodicidad y aparece la unidad que las constituye, que es la mol�cula de tropocol�geno, que tiene una longitud de 300 mm y un di�metro de 1.4 mm (o sea, que es una macromol�cula que ha sido medida). Cuando se neutraliza este medio �cido con �lcali, se puede observar que las fibrillas del tropocol�gano vuelven a agregarse muchas de ellas lado a lado, para reconstituir la fibrillas da col�geno nativa y vuelve a aparecer la periodicidad caracter�stica de las fibras col�genas. El peso molecular del tropocol�geno ha sido estimado entre 300.000 y 325.000. Cada mol�cula de tropocol�geno esta constituida por tres cadenas de polip�ptidos en cada una de las cuales hay alrededor de 1000 amino�cidos. La estructura de la triple h�lice del tropocol�geno es fundamental y es caracter�stica de esta prote�na fibrilar. Cuando existen defectos (incluso se conocen ciertos defectos gen�ticos) por los cuales existen d�ficit en algunos amino�cidos que constituyen la cadena de polip�tidos del col�geno, entonces la triple h�lice no se puede formar y en esos casos la mol�cula de tropcol�geno es defectuosa e incapaz de reconstituir la fibrilla de col�geno (o sea, no existe o no se forma el col�geno). Eso se ve en algunas enfermedades, algunas de origen hereditario y otras producidas por sustancias qu�micas, drogas, etc. Cuando se analiza ya la composici�n qu�mica de estas cadenas de polip�ptidos que constituyen el col�geno, se ve que los amino�cidos que conforman el col�geno tienen una distribuci�n bastante regular, que es lo que caracteriza a las prote�nas. Encontramos una estructura que se llama repetitiva en la secuencia de amino�cidos que se simboliza de esta manera: Gli � x � y � Gli � x � y � Gli � x- y - Gli
De modo que en s�ntesis lo que caracteriza al col�geno es esa secuencia repetitiva y la gran proporci�n que tiene de glicina, prolina e hidroxiprolina. La prolina y la hidroxiprolina constituyan juntas 22 % de todos los amino�cidos del col�geno. Se sabe que la hidroxiprolina desempe�a un papel fundamental y especial como elemento que estabiliza esta triple h�lice. Cuando hay defectos de la hidroxiprolina se traduce en la desorganizaci�n de la triple h�lice y por lo tanto de todo el col�geno. Finalmente, existen otros dos amino�cidos que se encuentran solamente en el col�geno, que son lisina y la hidroxilisina. Para terminar con esta parte de la anatom�a de la mol�cula del tropocol�geno, tenemos que hacer un peque�o agregado a esta disposici�n en triple h�lice. Las tres mol�culas est�n perfectamente entrelazadas a lo largo de toda la mol�cula de tropocol�geno menos en las puntas, aqu� se pierde la triple h�lice, de modo que podemos imaginar la mol�cula de tropocol�geno como una barra (un cilindro) y en las extremidades las tres mol�culas polip�ptidicas m�s desorganizadas y estas puntas son las que precisamente intervienen para formar uniones qu�micas con las mol�culas de tropocol�geno adyacentes. El tropocol�geno como tal se forma en el fibroblasto y sale de �l, pero la fibrilla de col�geno se forma s�lo por la agregaci�n ordenada de este tropocol�geno y esa agregaci�n ordenada se da tambi�n de una manera muy regular y espec�fica, que es lo que veremos ahora. Cada mol�cula de tropocol�geno la podemos representar de esta manera:
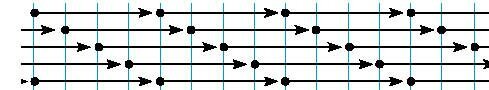
0 sea, una mol�cula que tiene 300 mm de longitud y que adem�s est� polarizada con dos extremidades diferentes. En un primer sentido, las mol�culas de tropocol�geno se ordenan a lo largo unas de otras, pero en la segunda hilera de mol�culas, en el col�geno nativo, hay una hilera que vendr� m�s de atr�s y as� sucesivamente se colocan desfasadas.
La cuarta mol�cula coincide con la primera. Esta disposici�n desfasada explica la aparici�n en el microscopio electr�nico cuando se utilizan colorantes, una serie de bandas transversales que resultan del alineamiento en sentido transversal de distintas partes de la mol�cula de tropocol�geno que est�n dispuestas de esta manera. Esta disposici�n ordenada de las macromol�culas explica tambi�n una propiedad f�sica y �ptica fundamental que es la birrefingencia, que es uno de los m�todos que se ha empleado para distinguir las distintas modalidades del col�geno. Revisi�n de los distintos tipos de col�geno:
|
<<<Regresar a T�cnica del Cuero>>>
Cueronet.com
Copyright � Cueronet.com
Reservados todos los derechos.